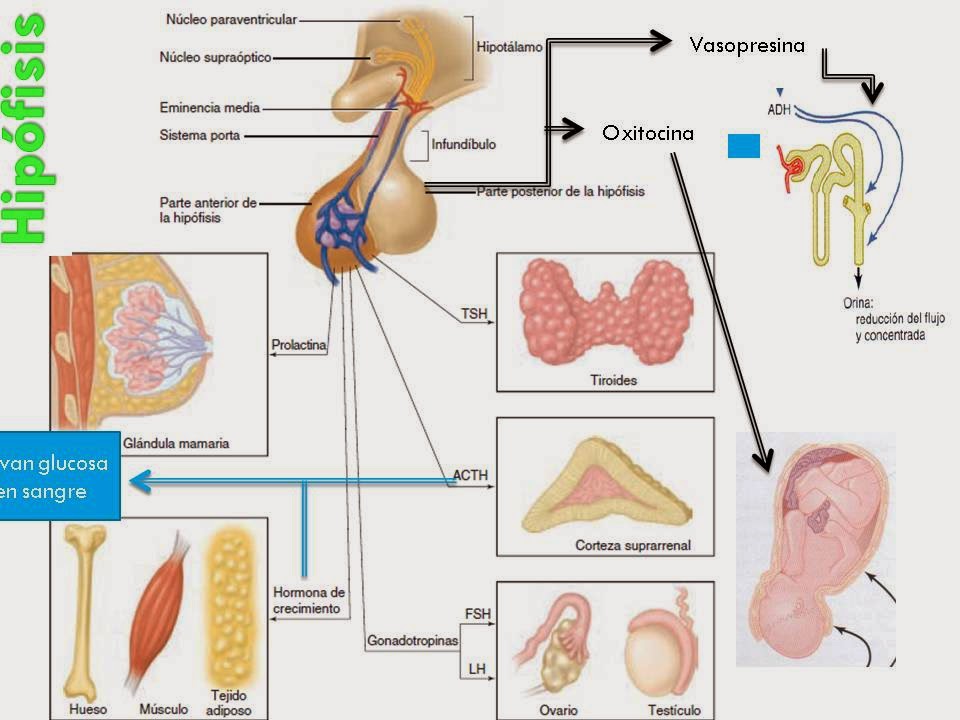
Eje Hipotálamo-Hipófisis Ovario
Cambios cíclicos en los ovarios:
Durante la fase folicular de los ovarios, que persiste desde el dia 1 hasta el 3, algunos de los foliculos primarios crecen, presentan vesículas, y se vuelven foliculos secundarios. Hacia el final de la fase folicular, un folículo de un ovario alcanza la madurez y se convierte en un folículo de Graaf.
Al crecer los foliculos, las células de la granulosa secretan un cantidad creciente de estradiol, que llega a su máxima concentración en la sangre dos dias antes de la ovulación, aproximadamente el día 12 del ciclo. La secreción del estradiol depende de la FSH secretada por la adenohipofisis. La FSH estimula la producción de receptores de FSH en las células granulosas. El estradiol también estimula la producción de nuevos receptores de FSH en los folículos. El estradiol aumenta la capacidad de la hipófisis para responder a la GnRH con un incremento de la secreción de LH. Como resultado de esta retroalimentación positiva, el estradiol sobre la hipófisis, hay un incremento de la secrecion de LH en la fase folicular tardia que culmina en un pico de secreción de LH.
El pico de secreción de LH comienza unas 24 hrs antes de la ovulación y alcanza su máxima intensidad 16 hrs antes de la ovulación. Durante este aumento de la secreción es cuando se detona la ovulación.

Después de la ovulación, el folículo vacío es estimulado por la LH para convertirse en una nueva estructura; el cuerpo luteo. El cuerpo luteo secreta estradiol y progesterona. La progesterona aumenta con rapidez hasta un grado máximo durante la fase lutea, aproximadamente una semana después de la ovulación. Las altas concentraciones de progesterona en combinación con el estradiol durante la fase lutea ejercen una retroalimentación negativa sobre la secrecion de FSH y LH. El cuerpo luteo producce inhibina durante la fase lutea, la cual ayuda a surprimir la secrecion o la accion de FSH. Esto sirve para retrasar el desarrollo de foliculos nuevos, de manera que la ovulación adicional normalmente no ocurre durante ese ciclo. Las concentraciones de estrogeno y progesterona descienden durante la fase lútea tardía, es alrededor del día 22.
Control Endocrino del ciclo ovárico
Cambios cíclicos en el endometrio
Fase Proliferativa: las cantidades crecientes de estradiol secretado por los foliculos en etapa de desarrollo estimula el crecimiento del estrato funcional del endometrio. El estradiol tambien estimula la producción de proteinas de recepetor para la progesterona en esta etapa, en preparación para la siguiente fase del ciclo.
Fase Secretora: ocurre cuando el ovario esta en su fase lútea. El incremento de secrecion de progesterona por el cuerpo luteo estimula el desarrollo de las glándulas uterinas. Las acciones combinadas de estradiol y progesterona engrosa el endometrio y las glándulas uterinas se congestionan con glucógeno durante la fase siguiente a la ovulación, preparado para aceptar y nutrir un embrión en caso de que ocurra la fecundación.
Fase Menstrual: ocurre como resultado del descenso de la secreción de hormonas ováricas durante la fase lútea tardía.
Eje Hipotálamo Hipófisis Testículos
Espermatogenia
Durante la formación del embrión, las células germinales primordiales emigran hacia los testículos y se convierten en células germinales inmaduras llamadas espermatogonias, comienzan a dividirse por mitosis a partir de la pubertad y continúan proliferando y diferenciándose a los estadios definitivos
de desarrollo para formar espermatozoides.
La espermatogenia tiene lugar en los tubulos seminiferos por estimulacion de las gonadotropas de la adenohipofisis, las espermatogonias emigran hacia la luz central del tubulo seminifero entre las celulas de Sertoli y ellas las rodea. Las espermatogonias que atreviesan la barrera y penetran en la capa de celulas Sertoli se modifican progresivamente y aumentan de tamaña para formar espermatocitos primarios grandes. Cada espermatocito primario se divide para formar dos espermatocitos secundarios, estos se dividen en espermatides. Aqui los 46 cromosomas del espermatocito se reparten, de manera que 23 cromosomas van a una espermatide y los otros 23 a la otra, que terminan conviertiendose en espermatozoides. Todo el periodo de espermatogenia, desde la espermatogonia hasta el espermatozoide, tiene una duración aproximadamente de 74 días.
Para mejor comprension del tema, te invito a ver este video :)